Αmphibians

Frogs, salamanders and caecilians display a stunning variety: some animals with tails and others without, some looking like snakes or lizards, others hopping on long hind legs, and with colors ranging from faded browns to bright blues, greens and reds.
Of 64,000 known species of vertebrates, only about 6,000 are amphibians. They are the smallest class of living vertebrates, all that remain of a group of animals that dominated prehistoric times, some of them even reaching the length of a medium-sized crocodile, that flourished several hundred million years ago. Frog and toad species make up the large majority of the known species. New amphibians are frequently discovered. In Madagascar, not long ago, a scientific survey found 106 species that had not been identified before. Herpetology or “the study of things that creep” is the branch of zoology that includes the study of amphibians but it also includes the study of reptiles.
Amphibians are an important group for study because they were the first back-boned, four-limbed creatures to walk the earth and also the group which later gave rise to reptiles/dinosaurs (as reptiles later gave rise to mammals and birds). Living amphibians are divided into three orders: Urodela (the salamanders, including newts and sirens), Anura (the frogs, including toads) and Gymnophiona (the legless caecilians).
The word “amphibian,” from the Greek amphibios, means a being with a double life – one that lives alternately on land and in water. Such a double life is the rule for Amphibia, but there are exceptions with some species permanently aquatic and others completely terrestrial. All are ectotherms, using environmental temperature to regulate body temperature.
No structure uniquely defines all amphibians (as feathers define birds for example) so a combination of characteristics must be considered. Further complicating any definition is the fact that living species have diverged significantly in the long history of the group from the primitive fossils and there are no fossil records about certain key features.

Evolution
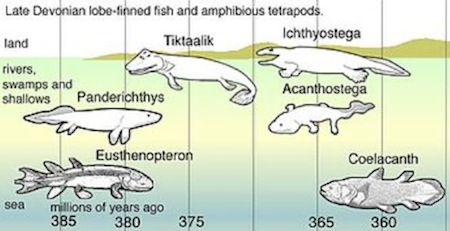
Some of the oldest known amphibians are Metaxygnathus, found in the freshwater beds of late Devonian deposits in Australia, and Ichthyostega and Acanthostega, found in similar beds in Greenland. All these fossils date from about 360 million years ago. Greenland may seem an unlikely place for any amphibian to have lived but its location and climate were very different during the Devonian period (410 – 345 million years ago), when it straddled the equator and lay within a moist and warm tropical region extending from present-day Australia through Asia to northeastern North America.
Until the early Jurassic period (about 190 million years ago), all of the earth’s land mass was united into a single supercontinent called Pangaea. Therefore, it is not surprising to find evidence that the earliest amphibians rapidly spread to now distant lands including Europe and eastern North America and, by the early Triassic (about 230 million years ago), even to Antarctica.
The most likely ancestors of these early Amphibia were lobe-finned fishes of the order Crossopterygii. Unlike other members of the fish class Osteichthyes, which had fins supported by cartilaginous rays, the fins of crossopterygians had bony elements comparable to those of the limbs of land vertebrates.
Some crossopterygians had lungs and some had internal nostril openings, so air could be taken into the lungs when the mouth was closed or when only the external nostrils were above water. Internal nostrils are characteristic of land vertebrates. In most fish the external nostrils serve only a sensory function – they lead to blind pockets not connected to the mouth cavity.

Ichthyostega, an early four-footed group, although unquestionably amphibian, retained a number of fish-like characteristics in the skeletal structure but the limb and girdle structure of ichthyostegids had already fully reached the early amphibian stage. This means that the earliest amphibian records (and land vertebrates) remain undiscovered and are still being sought.
How did the transition to land from the water come about? One classic explanation has been that the Devonian was a period of severe droughts. Fish with sufficiently strong fins could avoid stranding and death by crawling to available pools. According to this idea, land vertebrates could have evolved as a by-product of selection originally for increased agility in finding water, not land. Evidence casts doubt upon the scenario of periodic droughts however, and it seems likely that the Devonian was a time of relatively continuous moist environments, at least in tropical regions.
Possibly some of the features associated with the first amphibians actually developed in the aquatic environment. For example, the development of a functional neck and the separation of the skull from the pectoral girdle to accomplish this may have developed in proto-amphibians, permitting sudden sideways movement of the head to capture prey in water. Perhaps this change was a pre-adaptation later facilitating the capture of prey on land.
One or more of the following factors are believed to have led to the evolution of land vertebrates. During Devonian times aquatic environments, with their enormous diversity of fish and other organisms, contained many more competitors and predators than did the land, and land also may have been a safer place to deposit eggs and for juveniles to survive. The water of the warm Devonian swamps in which amphibians arose was probably poor in oxygen, especially in the shallows, but the fish ancestors of amphibians must have had lungs, as all of their living descendants do. Possibly these fish congregated in shallow waters, and ventured occasionally onto land. It might have been the more agile juveniles that did so, in order to exploit insect and other invertebrate food. Although this transition doubt-less occurred over a period of millions of years, there is no known fossil record of these stages. Some have even argued that the fish-to-amphibian transition occurred more than once, making amphibians, including modern species, members not of a single line, but of several that evolved independently.
In becoming terrestrial, amphibians overcame numerous challenges, although some changes could have occurred even in a shallow water habitat. On land, gravity became a key factor molding the development of the skeleton. Without the buoyancy of water, the body was suspended from the vertebral column which, in turn, had to be supported by the limbs and limb girdles. When the animal rested on the ground, a well-developed rib cage, as was present in Ichthyostega, prevented injury to internal organs. Other bones help distribute gravitational forces more evenly along the vertebral column. The skin of living amphibians, which is moistened by the secretions of numerous mucus glands, is not a passive outer layer but plays a vital and very active role in water balance, respiration and protection. It is highly permeable to water, especially in terrestrial species. Aquatic species have reduced permeability to offset the inflow of water by osmosis.

Although most amphibians are restricted to moist habitats, there are specializations that permit many species to live in otherwise inhospitable environments. For example, desert toads create an osmotic gradient across their skin by retaining urea in their urine, thus permitting water uptake from extremely dry soils. Most terrestrial frogs possess a patch of skin, rich in blood capillaries, in the pelvic region that allows uptake of water even from a thin surface film. Other frogs and a few salamanders form a cocoon of shed skin to reduce water loss, and some tree frogs reduce evaporative water loss by wiping fat-like skin secretions over the body surface.
On the other hand, loss of body water through the skin is used in some species as a method of temperature regulation through evaporative cooling. In most species the skin and surfaces in the mouth cavity also serve a respiratory function, since dissolved gases pass across them. The numerous members of one family of salamanders (Plethodontidae) have lost lungs altogether and depend entirely upon this method of gas exchange.
Practically nothing is known about the skin of the earliest amphibians. It is often assumed that they too had soft naked skin, but recent fossil evidence suggests instead that scales covered their undersides. Some types had bony scales on the upper surfaces.
After the appearance of the first amphibians in the late Devonian, a period of rapid evolution occurred resulting in an enormous diversity of amphibian types. However, by the end of the Triassic, about 155 million years later, nearly all of them had become extinct. Some were truly enormous in size. The largest, Mastodonsaurus, had a skull 49 inches (125cm) long and a length estimated at 13ft (4m). The largest currently living amphibians are the Asiatic representatives of the giant salamanders, which reach 67.5in (170cm).
Many species were aquatic and possessed gills, whereas others were adapted to land. Although many were heavy-bodied and lizard-like in build, there were some truly bizarre types including legless, eel-shaped species and others with extremely wide heads drawn back into peculiar horns.
These ancient amphibians were found on all land masses and were the dominant land animals of their day. Mammals and birds did not evolve until after most of these ancient amphibian types had become extinct, but the first reptiles evolved from amphibians very early in the Carboniferous period (about 345 million years ago). The earliest known reptiles resemble members of the amphibian order Anthracosauria.
As evidence of this close similarity, one anthracosaur (Seymouria) was long considered to be a primitive reptile. Recently, however, fossilized aquatic larvae were found, suggesting that seymouriamorphs did not lay shelled eggs. Nevertheless, the earliest reptiles were generally small and their rapid spread and evolution did not begin until late in the Permian (about 225 million years ago) at which time few of the ancient groups of amphibians still survived.

Modern Amphibians
In contrast to our knowledge about the origin of reptiles, the ancestry of modern amphibians is a puzzle, largely because there are no fossils linking the ancient Paleozoic types to any of the three living orders. The earliest known fossil (Triadobatrachus, of the early Triassic) is already frog-like in some of its features and the earliest true salamanders (from the late Triassic), true frogs (from the early Jurassic) and caecilians (from the early Tertiary, about 65 million years ago) are already as specialized, each in its own way, as modern ones.
The incomplete fossil record provides little help. It is curious why these animals were not readily fossilized, since even very small and fragile labyrinthodont larvae have been recovered. The reason may be ecological – the ancestors of living amphibians probably occupied very shallow waters or rushing mountain streams where the large species of ancient amphibians could not pursue them and these are places where fossilization is relatively rare. Without critical fossil material, guesses about evolution will have to rely on comparisons of the living species. For a long time, given the enormous differences between frogs and salamanders, it was believed that each had descended from different orders of Paleozoic amphibians. More recently, it was suggested that despite their different appearances, frogs, salamanders and caecilians have many basic features in common, particularly in the skin, ears, skull and teeth. The possibility that all of these features evolved independently is unlikely and it is more reasonable to assume a common origin. Therefore, many scientists place the three living groups in one subclass (Lissamphibia). But then newer evidence coming to light, particularly concerning the vertebral column, skull and jaw musculature, which raises anew the possibility that the living species arose from different orders of ancient stocks. Until critical fossil material from the late Paleozoic is found, the question is likely to remain unresolved.
Whatever their true origins, modern amphibians do share numerous features, in particular their highly permeable skin and their teeth but there has been a striking adaptive separation among them in terms of locomotion and reproduction.
Salamanders and caecilians swim like fish, with side-to-side sinusoidal movements. Frogs, on the other hand, swim (and jump) in a totally different way. The vertebral column has become progressively shortened, the hindmost vertebrae have fused into a single element, and the bones of the hind legs have become elongated. Frogs have rather inflexible bodies and swim by means of simultaneous thrusts of the legs.
Terrestrial salamanders move by means of lateral undulations, advancing diagonally opposite feet each time the body bends, some species use the tail as a fifth leg. Caecilians are legless and, except for a few completely aquatic species, live in burrows. Since the burrow walls greatly restrict lateral undulations, caecilians move by an alternating fold-and-extension progression in which only the vertebral column bends, producing momentary points of contact with the ground which allows extension of other parts of the body, superficially resembling the locomotion of an earthworm.
Reproduction
Amphibians exhibit the greatest diversity of reproductive modes of any vertebrate group. Fertilization can be external or internal. In the most primitive families (like the giant and Asiatic salamanders) it is external, the sperm being shed into the water near the eggs. However, most salamanders transfer sperm in small packets called spermatophores that are picked up by the female with her cloacal lips during courtship. The sperm can then be used at once or, in most species, stored in specialized glands in the cloaca for use during the following season. In two North American mole salamanders, the sperm merely activate the developmental process and do not contribute genetically.

With few exceptions, frogs fertilize externally. Usually sperm is deposited as the eggs are laid, with the male clasping the female with his forelegs. Some poison-arrow frog males fertilize the eggs after deposition. In some narrow-mouthed frogs the bodies are temporarily glued together, and in a few other species the male’s cloaca is held next to the female’s while sperm is transferred, so that fertilization is internal. but there is no intromittent organ. However. in the North American Tailed frog the tail is, in fact, an extension of the male’s cloaca which is inserted into the female’s cloaca to transfer sperm. All caecilians fertilize internally, the male using the cloaca as an intromittent organ. Most amphibian species lay eggs (they are oviparous) in fresh water or on land. Others are viviparous – the mother retains the eggs in her body and the embryos are nourished either by food stored in their own yolk sac (this is sometimes called “ovoviviparous” reproduction) or by materials obtained directly from the mother. Clutch size in frogs varies from species to species and ranges from a single egg to about 25,000. In salamanders, the number is generally no more than a few dozen. Fertilized eggs may be laid singly, in clusters, or in long strands, but are always enclosed in gelatinous envelopes. If laid in water (or near enough that hatched larvae can crawl or be swept into it by floods), the larvae have gills and lead an aquatic existence, eventually metamorphosing into miniature adults.
Amphibians use a variety of sites for laying eggs, including still or running water, mud basins constructed by the male, cavities beneath logs or stones, debris or burrows, leaves overhanging water, or the water-filled axils of plants. However, each species generally has one preferred site. Those that lay eggs on land typically have no free-living larval stage and undergo direct development; this is true in many tropical frogs and in virtually all terrestrial species of salamanders. In many frogs and salamanders the eggs are defended by one of the parents and in several species of frogs the eggs or tadpoles are carried.
Most amphibians giving birth to fully-developed young are ovoviviparous. In the Puerto Rican live-bearing frog, for example, only the embryo’s own yolk is utilized; the embryo’s tail is thin and rich in blood supply and may function in gas exchange. Several species of African live-bearing toads (genus Nectophrynoides) are ovoviviparous but in one, the West African live-bearing toad, the fetuses ingest a mucoprotein (uterine milk) secreted by the oviduct when the yolk supply is exhausted.

Two European species of salamanders are known to be viviparous. In the European Fire salamander the young are deposited as larvae into the water. In the Alpine salamander, a member of the same genus, only 1-4 young survive from as many as 60 fertilized ova. The survivors cannibalize their own siblings and they metamorphose before birth. The olm may also be viviparous under certain conditions. Although many caecilians lay eggs which are guarded by the mother, about half of the species are viviparous. After utilizing their own yolk, the large fetuses feed on uterine milk and also scrape material from the oviduct walls with their specialized teeth. Gas exchange occurs between the greatly-enlarged gills and the wall of the oviduct, but the gills are resorbed before birth. These are all of the truly live-bearing amphibians, so far as is known. They should not be confused with those instances of parental care in which parents only appear to give birth to fully-formed offspring.
The Amphibia are an astonishingly diverse class of vertebrates that has existed for at least 360 million years. Since at least the beginning of the Carboniferous period (about 340 million years ago) their lineage has been evolving independently. It would be wrong to think of modern amphibians merely as transitional form between fish and reptiles, except in a very general way. There is a tendency to regard living amphibians as evolutionary losers, partly because of their diminished numbers and small size. Instead, one should think of them as descendants of an ancient lineage that has been extraordinarily successful in exploiting an extremely wide range of habitats and life histories and which often constitutes a dominant element in natural communities.