Reptiles

The orders of reptiles are:
- Crocodilia (crocodiles, alligators, and caimans) – 23 species
- Sphenodontia (tuatara from New Zealand) – only 2 species
- Squamata (amphisbaenians (worm-lizards), snakes, and lizards) – about 7,900 species
- Testudines (tortoises and turtles) – about 300 species
Snakes, crocodiles, lizards – all animals that cause revulsion. Often they are thought of as cold, lurking animals, ancient relics of a group that is on its way out, superseded by birds and mammals. Their forerunners, the dinosaurs and other “prehistoric monsters,” are the best known of all extinct animals, with a firm place in popular fantasy. They are accused of being cumbersome and stupid. However, reptiles are in many respects much more successful than is often thought. In some habitats, notably deserts, they are a dominant group; the snakes have a recent evolutionary history of great diversification. Reptiles have one huge advantage over birds and mammals – being less dependent on maintaining a constant body temperature, they can survive on a fraction of the vast food input that birds and mammals require. They are thus able to exploit environments where food supplies are sparse or sporadic. The reptiles’ most obvious feature is their covering of dry, horny scales, which amphibians do not possess. They resemble primitive amphibians and birds in having only a single small bone in the ear, the columella or stapes, to conduct sound vibrations, and they have several bones in each side of the lower jaw. Mammals, on the other hand, have three small auditory bones and a single bone, the mandible, in the lower jaw, which corresponds to the reptilian dentary bone. Unlike birds and mammals, reptiles are mainly dependent upon external sources of heat such as the sun’s rays for maintaining their body temperature. They reproduce by laying shelled eggs on land or by bearing their young alive. They do not pass through an aquatic larval stage like most amphibians. Their embryos, like those of birds and mammals, are provided with special membranes, notably the amnion, which are of great importance in terrestrial reproduction.

Origins and Classification
The reptiles arose from amphibians of some kind, but the details of their early history are not clearly understood and current ideas about them are in a state of flux. Remains of the most primitive known reptiles have been found inside fossil tree stumps of the early Upper Carboniferous (about 315 million years ago). They were small terrestrial creatures superficially like lizards. It is likely that they were ancestral to most if not all the principal groups of later reptiles. However, it would seem that even at this ancient period the reptiles had already split into two main lineages, for there are less numerous remains from the same time of larger reptiles of a different type. These were the earliest members of the mammal-like line which ultimately led to the first tiny mammals at the end of the Triassic period (about 190 million years ago). The “cheek” or temporal region of the skull behind the orbits (eye sockets) has long been regarded as important in reptilian classification. A key feature distinguishing the various subclasses of reptiles is the number and nature of holes in this region. In the most primitive reptiles it presents an unbroken shell of bone without openings or “apses” (arched recesses). For this reason they are placed in the subclass Anapsida. In reptiles of the mammal-like line, a single opening has appeared with a bar of bone beneath it, a feature which may have been associated with changes in the attachments of the jaw muscles. These mammal-like reptiles belong to the subclass Synapsida.
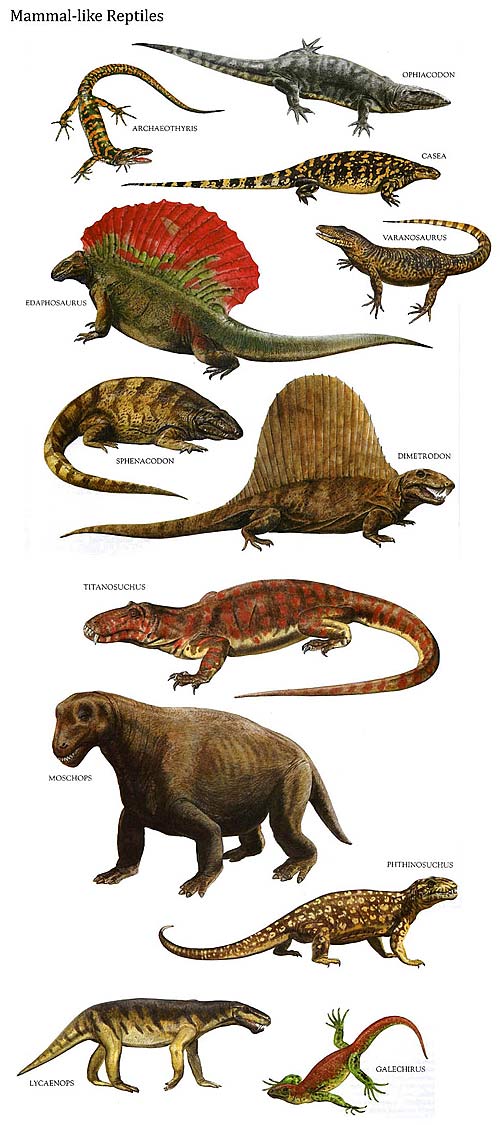
In Permian times (280-225 million years ago) another group of reptiles appeared, with two temporal openings on each side of the skull, each opening bounded below by a bony bar or “arch.” These belong to the subclass Diapsida which contains the majority of reptiles, both living and extinct. Two main lines of diapsids became numerous and diverse towards the end of the Triassic, when the mammal-like synapsids were disappearing. One line is called the Lepidosauria, not a helpful term since it just means “scaly reptiles.” It contains most living reptiles – the tuatara, conventionally placed in the order Rhynchocephalia (although this classification is due for revision), and the three suborders of the order Squamata —the lizards, the snakes, and the worm lizards, a group of worm-like burrowing reptiles. The tuatara retains the typical skull structure of the early diapsids with two distinct temporal openings. In many ways it can be regarded as a “living fossil.” Close relatives occurred in the Triassic. In “typical” lizards the upper temporal opening is surrounded by bone but the bar beneath the lower one has disappeared. The earliest lizards occurred in the Triassic, but snakes do not appear until the Lower Cretaceous (about 135 million years ago). They are the most recently evolved of all the bigger groups of reptiles, and in terms of numbers of families and species they have been spectacularly successful. They most probably arose from lizards, perhaps from unknown burrowing types which must, however, have been very different from the various kinds of burrowing limbless lizards alive today. The other principal diapsid line was the Archosauria which became immensely successful during the Jurassic and Cretaceous periods (between about 190 and 65 million years ago). It is these “ruling reptiles” that draw the human imagination to the Jurassic and the Cretaceous, above all other periods in the Age of Reptiles. The most spectacular archosaurs were the dinosaurs, mostly large or enormous creatures, although a few were little bigger than a pheasant. There were two distinct orders of them, the Saurischia, in which the hip bones were of the typical reptilian pattern, and the Ornithischia, “bird-hipped” forms. The Saurischia contained all the carnivorous bipedal types, culminating in Tyrannosaurus, with an overall length of 40 ft (12m), and also immense quadrupedal herbivores, the brontosaurs, with tiny heads and long necks and tails. One of these, Brachiosaurus, with a length of 75ft (23m) and an estimated weight of 50 tons, was one of the heaviest land animals that has ever existed, although incomplete remains of an even larger relative have been reported. The Ornithischia also contained both bipedal and quadrupedal types, but all were herbivorous. The bipeds include the well-known Iguanodon with a spur on its thumb, and the duck-billed dinosaurs, some of which possessed curious crests of disputed function on the tops of their heads. Among the ornithischian quadrupeds were the lumbering Stegosaurus with huge plates of bone down its back, and the rhinoceros-like horned dinosaurs such as Triceratops. Some workers believe that the birds were descended from some kind of small primitive dinosaur, and the first known bird, Archaeopteryx, might well have been classified as a dinosaur if it were not for its feathers. Other successful archosaurian groups were the flying reptiles (Pterosauria) and the Crocodylia, which originated like the dinosaurs and most of the other, better-known groups of reptiles in the Triassic. The flying reptiles were similar to birds in certain features, such as the keel on the sternum for the attachment of the flight muscles and the presence of air spaces in many bones to reduce their weight. However, the origin of the two groups seems to have been quite independent and the pterosaurs flew on wings quite unlike those of birds, being membranous and supported by the elongated bones of the fourth finger. The Crocodylia, beautifully adapted for an amphibious predatory mode of life, are the only surviving archosaurs and their continued existence is threatened by the leather trade. A still unresolved problem of great geological and biological interest is why the crocodiles should have survived the mass extinction of reptiles at the end of the Cretaceous, when the dinosaurs and other spectacular groups became extinct. There are two further groups of reptiles which do not fit easily into either the synapsid or the diapsid pattern. These are the great Jurassic marine reptiles, the fishlike ichthyosaurs, and the plesiosaurs with long necks and chunky bodies. Very little is known of their origins and there is no evidence that they were closely related. However, they both possessed a single temporal opening high up on the side of the skull and are tentatively placed in the subclass Euryapsida. Finally there is the order Chelonia or Testudines, the tortoises and turtles, in some ways both the oddest and the most familiar of reptiles. They appear almost “fully fashioned” in the Triassic and virtually nothing is known of their origins. Unique features of their anatomy, however, suggest that they branched off from the primitive reptilian stock at a very early stage in evolution. In a few forms, such as the sea turtles, the skull seems to resemble the primitive anapsid pattern, but in many others certain bones appear to have been “eaten away” or emarginated from behind to produce a kind of temporal opening anatomically different from the one in synapsids, diapsids or euryapsids. They are provisionally placed in the subclass Anapsida, together with the most primitive known reptiles.

The Skin
The skin is a fascinating organ with many functions. Besides acting as a barrier between the deeper tissues of the animal and the outside world, it can have a role in defense, concealment, mating and locomotion. In reptiles, as in other vertebrates, it consists of two principal layers, the epidermis on the outside and the dermis beneath. The epidermis is itself subdivided into various layers, the outer one composed of the horny material called keratin. The scales of a reptile are localized thickenings of this keratin layer, connected by hinges of thinner material and often folded back so that they overlap each other. Unlike the scales of fish they are not separate, detachable structures but parts of a continuous epidermal sheet. Their precise numbers and patterns on different parts of the head and body are of great value in reptilian classification, especially in the distinction between different species. Periodically the keratinous layer of the epidermis is shed and replaced through the activity of the deeper layers of cells. The keratin may come away piecemeal or in large flakes. In snakes, however, it is often shed as a single slough which is peeled off inside out after the snake has rubbed it through at the snout. In these reptiles the old keratin layer on the surface is not shed until a new one has completely formed beneath it. Then a translucent zone of cleavage appears between the old and new layers so that they can easily become separated. The brille, the spectacle-like eye-covering of a snake, formed from modified and fused eyelids, becomes blue and opaque some time before the animal is due to shed. Just before this happens, however, it becomes clear again and this clearing seems to coincide with the establishment of the cleavage zone. Skin-shedding in snakes may occur several times a year, being more frequent in young than in old animals. The process is influenced by the activity of the thyroid gland. The dermis consists mainly of connective tissue and contains many blood vessels and nerves; it does not participate in the skin-shedding process. In some reptiles, including crocodilians and many lizards, it contains plates of bone called osteoderms which lie beneath and reinforce the horny epidermal scales. In the slowworm, and certain other lizards, these osteoderms form a flexible bony covering for the body. The huge bony frills and plates of dinosaurs such as Triceratops and Stegosaurus were structures of this type, and probably had a horny covering which did not become fossilized. Both epidermis and dermis participate in that remarkable structure, the shell of a turtle. The horny scutes on the surface are keratinous epidermal formations with a layer of living cells beneath them, while the deeper, thicker shell layer is formed of bony dermal plates. In reptiles the dermis also contains the majority of pigment cells. Many of these, the melanophores, contain black pigment, but there may also be white, yellow and red pigment cells. The dispersal or concentration of the pigment within the melanophores, and the optical effects of this when seen through the colored cells, are responsible for the changes in color for which chameleons and certain other lizards are noted. Color change in reptiles may be brought about by nervous activity or by the hormonal secretions of glands such as the pituitary, or by a combination of both; in chameleons the action of the nerves seems to be the dominant factor. The reptilian skin contains comparatively few glands, unlike that of many fish, amphibians and mammals. However, crocodilians possess a pair of glands beneath the throat which produce a musky secretion; this may play some part in sexual behavior. Some freshwater turtles have glands in the chin or the hind-limb pockets; the musk turtles these produce a powerful smell. A few otherwise non-poisonous snakes have glands beneath the scales of the neck or back which secrete an irritant material – a defense against predators and some role in courtship behavior have been suggested as possible functions. In some geckos of the genus Diplodactylus there are a series of large glands beneath the scales of the tail. When the lizard is threatened they squirt out filaments of sticky material which may deter such enemies as large spiders. Many lizards possess a series of curious gland-like structures on the insides of the thighs and sometimes in front of the cloaca. The function of these has been much debated; they seem to be related to mating since in male lacertid lizards they regress after castration. The most likely suggestion is that their secretions may assist in species or sexual recognition.

The Skeleton
Apart from its diapsid skull, the skeleton of the tuatara probably shows the greatest living resemblance to skeletons of primitive fossil reptiles. The other reptilian groups have departed in varying degrees from this primitive pattern, and such departures are usually in the nature of adaptations to special modes of life. The loss of the lower temporal bar in lizards has been associated with the development of hinges at various points in the skull, allowing increased mobility of the jaws (kinesis). Jaw mobility has been carried even further in snakes, partly because of the loss of any firm union between the two sides of the lower jaw. This is an adaptation to eating large prey. The reduction or loss of the limbs in some lizards and nearly all snakes and an increase in the number of vertebrae is a dramatic adaptation to a special lifestyle – one that has been extraordinarily successful, enabling the snake to “outswim the fish and outclimb the monkey,” as has been written with some exaggeration. The remarkable position of the limb girdles of tortoises and turtles, which are situated within rather than outside the ribs, is another striking specialization associated with the incorporation of the ribs into the dermal plates of the shell. The reptilian skeleton contains a number of bones which are not found in mammals, at least as separate elements. Some of these, such as the parasphenoid and the supratemporal, occur in the skull. There is also a midline bone, the interclavicle, in the shoulder girdle of most reptiles; among mammals it occurs only in the egg-laying monotremes. The tuatara, lizards and turtles have, like birds, a series of small bony plates, the scleral ossicles, in the eye which assist in focusing. In the tuatara, crocodilians and some extinct reptiles, the belly wall is reinforced by a series of abdominal ribs or gastralia. Many lizards have a system of cartilaginous bars called the parasternum in much the same position. Secondary centers of ossification (bony epiphyses) are lacking in the growing bones of most modern reptiles, although they do occur in many lizards. In mammals such as man the fusion of these with the main part of the bone in early adult life sets a limit to the maximum size attained. The absence of bony epiphyses is perhaps one of the reasons why certain reptiles such as pythons, crocodiles and giant tortoises may be able to grow throughout much of their lives, albeit at a diminishing rate. If an individual is lucky enough to avoid life’s hazards it may eventually become a giant, well above the usual size range of its species. However, many reptiles including the smaller lizards and turtles normally stop growing when they have reached a more or less definite size. Moreover, reptiles do not lose their teeth when they get old in the way that mammals do. Most reptiles have continuous, lifelong tooth replacement, while turtles, which did away with their teeth at an early stage in their history, have evolved a persistently growing horny beak. This removes a constraint on longevity and therefore also on the ultimate potential size attainable by a species.

Reproduction
Reproduction In most species of reptile the two sexes differ to some extent in adult size, shape or color. The males of many lizards, land tortoises and all crocodilians are bigger than the females, whereas in most snakes and some aquatic turtles it is the females that are bigger. Differences between the sexes are particularly striking among the agamid and iguanid families, lizards in which the eyes are the predominant sense organs. The males tend to be more brightly colored, especially in the breeding season, and in some species they possess erectile crests and throat fans which play a part in courtship and territorial display. All reptiles practice internal fertilization, the sperm being introduced directly into the female’s cloaca, the common opening which transmits both excretory products and eggs or sperm. In the tuatara fertilization is achieved by cloacal contact, but in all other living reptiles the males have special organs of insemination. In turtles and tortoises and the crocodilians there is a single penis. Lizards and snakes have evolved paired organs called hemipenes, only one is used at a time. The females of some snakes and some turtles and tortoises have the ability to store sperm in the reproductive tract and instances of isolated females laying fertile eggs after a year or more in captivity have been observed. Breeding is greatly influenced by environmental factors such as temperature and the duration of daylight. Some tropical species may breed at intervals throughout the year, but in the majority, breeding takes place only once, or perhaps twice a year. Indeed, among those which live in relatively severe climates such as the adder in northern Europe an individual female may breed only once every two years, or even less often. Sexual reproduction is the general rule among vertebrates but it has been discovered that some populations of rock lizards in the Caucasus region consist entirely of females and reproduce by parthenogenesis or “virgin birth.” This also happens in certain whiptail lizards in North America. In this method of reproduction, all members of the population are genetically identical, all descended from a single founding female. The low level of genetic variability in such a species both confines it to a limited geographical area and probably limits its future in evolutionary time, because of its reduced ability (compared with a sexual species) to adapt to changing environmental conditions. Reptiles are the “highest” animal group in which parthenogenesis normally occurs. Most modern reptiles lay eggs, which, unlike amphibians’ eggs, are resistant to drying out. Fossilized eggs of dinosaurs and more primitive extinct reptiles have been discovered. These eggs possess a shell which may be pliable or parchment-like in texture, as in many lizards, snakes and aquatic chelonians, or hard and well calcified, as in tortoises, crocodilians and many geckos. Hatching from the egg is facilitated by the presence in young lizards and snakes of a sharp, forward-pointing egg-tooth which is later shed in the tuatara, tortoises and turtles, crocodilians and birds a horny out-growth performs the same function. The replacement of aquatic amphibian eggs by the shelled amniote egg must have played a vital part in liberating the reptiles from their watery ancestral habitat and fitting them to become the first successful vertebrate colonists of the land. The eggs of reptiles are usually laid in holes, among rotting vegetation or buried in the soil. Sea turtles dig nests in sand on beaches and lay large clutches of up to 100 or more eggs. They resort to traditional nesting beaches, and Green turtles may migrate hundreds of miles to reach them. Some crocodilians such as the Nile crocodile also dig nests in the sand, but others such as the American alligator construct more elaborate nests out of piled-up vegetation. The rate of embryonic development is accelerated by warmth. It has been dis-covered that in certain turtles and tortoises and some crocodilians the sex of the hatchling depends on the prevailing temperature during a critical period of incubation. Thus, in the American alligator temperatures of less than 86°F (30°C) between the 7th and 21st days of incubation produce all females whereas those of more than 93°F (34°C) produce all males. It has been suggested that the possible occurrence of a similar method of sex determination in dinosaurs might have contributed to their extinction. If temperatures increased or decreased markedly at the end of the Cretaceous, a unisex population might have resulted. But why were the crocodiles apparently not affected? The majority of reptiles abandon their eggs after laying but in certain lizards, for example in some skinks, and also in a few snakes such as cobras, the female remains with her eggs and may try to drive intruders from the nest. Attendance by the male has also been reported in cobras. Female pythons remain coiled round their eggs for up to several weeks and in some species the egg temperature may be actively raised: this incubation may be assisted by muscular contractions of the mother’s body. During recent years remarkably elaborate forms of parental care have been observed in crocodilians. A very substantial number of lizards and snakes have forsaken egg-laying and gone over to live-bearing, the young escaping from their membranes at or shortly after birth. In such reptiles the eggshell is lost or reduced, and in many species a kind of placenta is developed from the fused chorion and allantois and from the yolk sac. This allows exchange of some waste products and nutritional substances, as well as water and gases, between the embryo and the mother. Most viviparous species are “ovoviviparous” – the yolk remains large and is still the principal source of embryonic food, but in a few with particularly well-developed placentae, the yolk is reduced. Most sea snakes have become viviparous, and therefore do not have to come on land to lay their eggs, as the turtles have to do. This method of reproduction is also prevalent in reptiles which live under very severe climatic conditions, as in high latitudes or altitudes. It is found in three out of the six species of British reptiles. Under such conditions viviparity seems to be advantageous in enabling the mother to act as a mobile incubator, able to seek out sources of warmth which are optimal for the development of her embryos.

